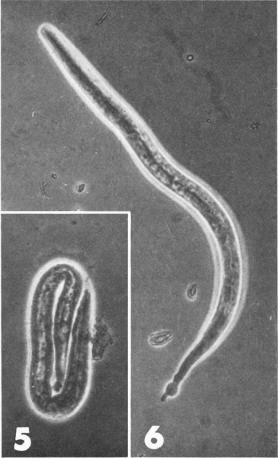
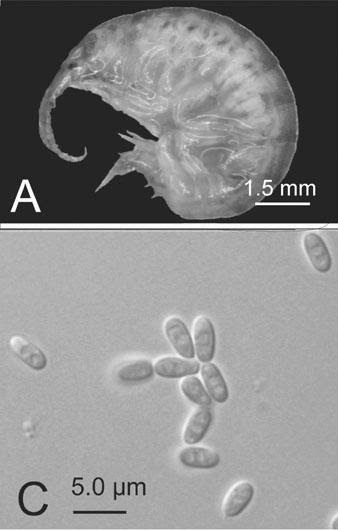
Hornets can put fear into the minds of many people, but today meet the parasite that the hornets fear (if they are capable of fear). Sphaerularia vespae is a parasitic nematode that infects the Japanese yellow hornet (Vespa simillima) and as far as infection goes, this one is quite a doozy. It specifically invade and resides in the gaster (abdomen) of female hornets where it grows and develop. The nematode ends up sterilising the host (much like other parasiticcastrators we have featured on this blog), turning her into a cozy nursery for baby worms. But a new study has shown that they are capable of doing more than just castrate the hornet.
In a previous study, a group of scientists noticed that the majority of overwintered hornet queens caught in bait traps were infected with S. vespae, so there is something about these nematode-infected hornets which seems to make them more likely to end up in those traps. During autumn/fall, queen hornets fortunate enough not to be infected with S. vespae would visit and poke around various nooks and crannies (usually decayed logs) in the forest to find a spot to hibernate. When the hornet find a place she likes, she will start excavating a hibernacula ( a place to hibernate) and line it with plant fibres that serve as nesting material. But queens that are parasitised and sterilised by S. vespae start visiting decaying logs much earlier during early to mid-summer.
A team of scientists in Japan decided to find out just what those infected queens are up to. For three months between May and August, they made regular weekly visits to a predesignated sites in a forest at the foot of Mount Moiwa and set up a video cameras to observe the decayed logs in the morning and afternoon.
They saw that unlike other hornets, the nematode-infected queens never dig nor gather nesting material. They simply crawl inside a decayed log, hang out for a while, then fly off. That is because they have become sterilised couriers that visited potential hibernation sites only to drop off a special package in the form of S. vespae juveniles. A quarter of the infect queens they saw landing on decayed logs offloaded some nematodes (there were some hornets that moved out of sight so the scientist couldn't see what they were up to). But in addition to those observations, the scientists also captured some hornet queens and brought them back to the laboratory for further examination. They kept them in vials and noticed that over two-third of the infected hornets ended up releasing juvenile worms.
When they dissected hornets to see how many of them were infected and to check the developmental stage of their parasites, they found a seasonal pattern to the infections. Queens caught during May and June were mostly infected with fully-mature female worms and their eggs, while queens caught between July and throughout August were filled with juvenile worms that were ready to disembark and infect a new host - which just so happen to be the period when parasitised queens start making regular visits to potential hibernation sites.
So that is S. vespae's game - use the hornet as a mobile incubator/nursery, fly her around during summer to scope out the best pieces of real estate around the forest, then drop off a bundle of worms that can lie in wait like a booby-trap for an uninfected hornet queen to come along and settle in for winter. To complete its life cycle, S. vespae simply take advantage of a preexisting behaviour (seeking out hibernation sites) from the host's repertoire, and "switch it on" at a different time of year to fit the developmental schedule of the parasite's own offspring. Parasite manipulation isn't necessarily about teaching an old host new tricks, but to get the host to perform the tricks that it already knows in a brand new context.
Reference:
Sayama, K., Kosaka, H., & Makino, S. (2013) Release of juvenile nematodes at hibernation sites by overwintered queens of the hornet Vespa simillima. Insectes Sociaux60: 383-388.
 |
| Photo of a queen hornet (from Fig. 2 of the paper) |
A team of scientists in Japan decided to find out just what those infected queens are up to. For three months between May and August, they made regular weekly visits to a predesignated sites in a forest at the foot of Mount Moiwa and set up a video cameras to observe the decayed logs in the morning and afternoon.
 |
| Photo of a hornet releasing some S. vespae juveniles (from Fig. 2 of the paper) |
When they dissected hornets to see how many of them were infected and to check the developmental stage of their parasites, they found a seasonal pattern to the infections. Queens caught during May and June were mostly infected with fully-mature female worms and their eggs, while queens caught between July and throughout August were filled with juvenile worms that were ready to disembark and infect a new host - which just so happen to be the period when parasitised queens start making regular visits to potential hibernation sites.
So that is S. vespae's game - use the hornet as a mobile incubator/nursery, fly her around during summer to scope out the best pieces of real estate around the forest, then drop off a bundle of worms that can lie in wait like a booby-trap for an uninfected hornet queen to come along and settle in for winter. To complete its life cycle, S. vespae simply take advantage of a preexisting behaviour (seeking out hibernation sites) from the host's repertoire, and "switch it on" at a different time of year to fit the developmental schedule of the parasite's own offspring. Parasite manipulation isn't necessarily about teaching an old host new tricks, but to get the host to perform the tricks that it already knows in a brand new context.
Reference:
Sayama, K., Kosaka, H., & Makino, S. (2013) Release of juvenile nematodes at hibernation sites by overwintered queens of the hornet Vespa simillima. Insectes Sociaux60: 383-388.